헤모글로빈 (Hb) 에 대해 포괄적으로 말할 수 있으려면 헤모글로빈 과 매우 유사하지만 훨씬 더 단순한 미오 글 로비 인 (Mb) 을 먼저 다루는 것이 유용합니다. 헤모글로빈과 미오글로빈 사이에는 혈연 관계가 엄격합니다. 둘 다 접합 된 단백질이고 그 보체 그룹 (비 단백질 부분)은 헴 그룹입니다.
Myoglobin은 약 150 개의 아미노산 (유기체에 따라 다름)의 단일 사슬로 구성된 구형 단백질이며 분자량은 약 18 Kd입니다.
언급 한 바와 같이, 그것은 단백질의 소수성 (또는 친 유성) 부분에 삽입되는 헴 군 (heme group)을 갖추고 있으며, 섬유 성 단백질의 α 나선 구조에 기인하는 접힘으로 구성된다.
미오글로빈은 주로 α-helix의 분절로 구성되어 있으며 8 개 존재하며 거의 극성이없는 잔기 (류신, 발린, 메티오닌, 페닐알라닌)로 구성되어 있으며 극성 잔기는 실질적으로 존재하지 않습니다 (아스파르트 산, 글루탐산, 라이신 및 아르기닌); 유일한 극성 잔기는 두 개의 히스티딘으로 헴 그룹에 대한 산소의 공격에 기본적인 역할을합니다.
헴 그룹은 발색단 그룹 (보이는에서 흡수)이며, myoglobin의 기능 그룹입니다.
또한보십시오 : 당화 된 헤모글로빈 - 소변에있는 헤모글로빈
약간의 화학 작용
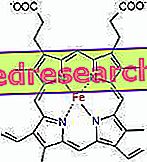
프로토 포르피린과 철 사이의 연결은 중심 원자 (또는 이온)가 산화 수 (전하)보다 높은 수의 다른 화학 종과 결합을 형성하는 화학 화합물 인 이른바 배위 화합물의 전형적인 결합이다. 헴의 경우 이러한 결합은 가역적이며 약합니다.
철의 배위 수 (배위 결합 수)는 6입니다. 철 주위에 결합 전자를 공유하는 6 개의 분자가있을 수 있습니다.
배위 화합물을 형성하기 위해서는 올바른 방향성을 가진 두 개의 궤도가 필요하다 : 하나는 전자를 "사고"다른 하나는 그것들을 기증 할 수있다.
헴에서 철은 proto-porphyrin 고리의 중앙에있는 네 개의 질소 원자와 근위 히스티딘 질소가있는 다섯 번째 결합과 네 개의 평면 결합을 형성합니다. 철은 여섯 번째 자유 배위 결합을 가지며 산소에 결합 할 수 있습니다.
철이 자유 이온 형태로 존재할 때, d- 형 궤도는 모두 동일한 에너지를 갖는다. 미오글로빈에서 철 이온은 프로토 포르피린과 히스티딘에 결합한다. 이들 종은 철의 궤도를 자기 적으로 방해한다. 섭동의 범위는 그들의 공간적 성향과 방해하는 종의 그것에 따라 다양한 궤도에 대해 다를 것이다. 궤도의 총 에너지가 일정하게 돌아 가야하기 때문에, 섭동은 여러 궤도 사이의 에너지 분리를 유발합니다. 일부 궤도에 의해 획득 된 에너지는 다른 궤도에 의해 손실 된 에너지와 같습니다.
궤도 사이에서 발생하는 분리가 그다지 크지 않다면 높은 회전 전자 배열이 바람직합니다. 결합 전자는 몇 가지 가능한 하위 수준 (최대 다중도)에서 병렬로 배열하려고합니다. 반면에 섭동이 매우 강하고 궤도 사이에 큰 간격이 있다면 낮은 에너지 궤도 (낮은 회전 속도)에서 결합 전자를 짝을 이루는 것이 더 편리 할 수 있습니다.
철이 산소에 결합하면 분자는 낮은 회전 배열을 가지지 만 철은 6 번째 자유 배위 결합을 가지면 분자는 높은 회전을 갖습니다.
미오글로빈의 스펙트럼 분석을 통해 스핀의 차이로 인해 결합 산소 (MbO2)가 있는지 여부 (Mb)를 알 수 있습니다.
Myoglobin은 근육의 전형적인 단백질 이지만 (근육에서 발견되는 것은 아닙니다).
미오글로빈은 정자 고래에서 추출되어 대량으로 존재하고 정제됩니다.
고래류는 인간과 같은 호흡을합니다. 호흡 과정을 통해 폐를 흡수해야합니다. 향유 고래는 산소를 축적 할 수있는 근육에 가능한 한 많은 산소를 운반해야한다. 근육에 존재하는 미오글로빈에 산소를 결합시킨다. 고아가 산소를 필요로하기 때문에 고래가 잠겨있을 때 산소가 천천히 방출됩니다. 정자 고래가 흡수 할 수있는 산소의 양이 많을수록 다이빙 중에 더 많은 산소가 이용 가능합니다.

고기를 더 적거나 적게 만드는 것은 hemoproteins의 함량입니다 (고기를 빨간색으로 만드는 것은 heme입니다).
헤모글로빈 은 myoglobin과 많은 구조적 유추를 가지고 있으며 분자 산소를 가역적으로 결합 할 수 있습니다. 그러나 미오글로빈은 일반적으로 근육과 말초 조직에 국한되어 있지만 헤모글로빈은 혈액의 40 %를 차지하는 적혈구 또는 적혈구에서 발견됩니다 (즉, 그들은 실제 세포가 아닙니다.
헤모글로빈 은 4 량체의 폴리 펩타이드 사슬로 구성되어 있으며, 각각 4 개의 폴리 펩타이드 사슬 (heme group)과 2 개의 2 개의 사슬 (인간에서는 2 개의 알파 사슬과 2 개의 베타 사슬이 있음)로 구성되어 있습니다.
헤모글로빈 의 주요 기능은 산소의 수송이다 . 헤모글로빈이 관련된 혈액의 또 다른 기능은 물질을 조직으로 운반하는 것입니다.
폐 (산소가 풍부한)에서 조직까지의 경로에서 헤모글로빈은 산소를 운반하며 (동시에 다른 물질도 조직에 도달 함) 역방향 경로에서는 조직에서 수집 한 폐기물, 특히 신진 대사에서 생성 된 이산화탄소를 운반합니다.
인간의 발달에는 일정 기간 동안 만 나타나는 유전자가 있습니다. 이런 이유로 우리는 다른 헤모글로빈을 가지고 있습니다 : 성인 남성의 태아, 배아.
이 서로 다른 헤모글로빈을 구성하는 사슬은 구조가 다르지만 유사성을 통해 수행하는 기능은 거의 동일합니다.
몇 가지 다른 사슬의 존재에 대한 설명은 다음과 같습니다 : 유기체의 진화 과정에서, 헤모글로빈은 또한 결핍 지역이 풍부한 지역으로부터의 산소 수송을 전문으로 발전 시켰습니다. 진화 사슬의 시작에서 헤모글로빈은 작은 유기체에서 산소를 운반했다. 진화 과정에서 유기체는 더 큰 차원에 이르렀습니다. 따라서 헤모글로빈은 부유 한 지점에서 더 멀리 떨어진 곳으로 산소를 운반 할 수있게 변경되었습니다. 이를 위해 헤모글로빈을 구성하는 사슬의 새로운 구조가 진화 과정에서 체계화되었다.
Myoglobin은 겸손한 압력에서도 산소를 결합합니다. 말초 조직에서 약 30 mmHg의 압력 (PO2)이 발생합니다.이 압력에서 미오글로빈은 산소를 방출하지 않으므로 산소 운반체로서 효과적이지 않습니다. 한편, 헤모글로빈은보다 탄력적 인 행동을합니다. 즉, 압력이 낮아지면 고압에서 산소를 묶어 방출합니다.
단백질이 기능적으로 활성 인 경우, 단백질의 모양을 약간 바꿀 수 있습니다. 예를 들어 산소화 된 미오글로빈은 비 산소화 된 미오글로빈과는 다른 형태를 가지며이 돌연변이는 인접한 미오글로빈과는 다른 형태를 갖습니다.
담론은 헤모글로빈과 같은 관련 단백질의 경우 다르다 : 사슬이 산소화되었을 때 그것의 모양을 변화 시키도록 유도되지만이 변형은 3 차원이므로 다른 4 사슬도 영향을 받는다. 사슬이 서로 연관되어 있다는 사실은 하나의 수정이 다른 이웃 사물에도 영향을 미친다 고 생각하게합니다. 사슬이 산소가 공급되면 테트라 메터의 다른 사슬은 산소에 대한 "덜 적대적인 태도"를 취하게됩니다. 사슬이 산소화되면 체인이 산소가 감소하는 어려움이 줄어 듭니다. 동일한 주장이 탈산 소화에 유효합니다.
탈 옥시 헤모글로빈의 4 차 구조는 T (시제) 형태로 불리는 반면, 옥시 헤모글로빈의 구조는 R 형태 (방출 됨)로 불린다. 스트레칭 상태에서 산성 아미노산과 염기성 아미노산 사이에 다소 강한 정전기적인 상호 작용이있어 디 옥시 헤모글로빈의 단단한 구조로 이어진다 (이것이 왜 시제 형태인가?) 반면 산소가 결합되면 이들의 실체 상호 작용이 감소합니다 (이것이 "공개 된 양식"인 이유입니다). 또한, 산소가 없을 때, 히스티딘 전하 (구조 참조)는 아스파르트 산의 반대 전하에 의해 안정화되는 반면, 산소 존재 하에서는 단백질이 양성자를 잃는 경향이있다; 이 모든 것은 산화 된 헤모글로빈이 탈 산소 된 헤모글로빈보다 강한 산인 보아 효과 라는 것을 의미합니다.
산도에 따라 헤모글로빈은 산소에 쉽게 결합한다. 산성 환경에서 헤모글로빈은 산소를보다 쉽게 방출한다 (긴장된 형태는 안정하다). 반면에 기본적인 환경에서는 산소와의 결합이 강해.

각 헤모글로빈은 들어오는 산소 1 몰당 0.7 개의 양성자를 방출합니다 (O2).
Bohr 효과 는 헤모글로빈이 산소 운반 능력을 향상시킬 수있게합니다.
폐에서 조직으로의 여행을 만드는 헤모글로빈은 압력, pH 및 온도에 따라 균형을 이루어야합니다.
우리 는 온도 의 영향을 봅니다.
폐의 폐포의 온도는 외부 온도보다 약 1 ~ 1.5 ℃ 낮지 만 근육의 온도는 약 36.5 ~ 37 ℃입니다. 온도가 증가함에 따라 포화 인자는 감소합니다 (동일한 압력으로). 이는 운동 에너지가 증가하고 해리가 선호되기 때문에 발생합니다.

헤모글로빈이 산소에 결합하는 능력에 영향을 줄 수있는 다른 요인들도 있는데, 그 중 하나 는 2, 3 bisphosphoglycerate 의 농도입니다 .
2, 3 bisphosphoglycerate는 4-5mM의 농도로 적혈구에 존재하는 대사 작용제입니다 (신체의 다른 부위에서는 그렇지 않습니다).
생리 학적 pH에서, 2, 3 비스 포스 포 글리세 레이트는 탈 양자화되고 여기에 5 개의 음성 전하가있다; 헤모글로빈의 두 베타 사슬 사이에 쐐기 모양으로되어 있는데, 이 사슬은 양전하가 고농축이기 때문입니다. 베타 사슬과 비스 포스 포 글리세 레이트 사슬 간의 정전 기적 상호 작용은 시스템에 일정한 강성을 부여합니다 : 산소에 대한 친화력이 거의없는 시제 구조가 얻어집니다. 산소가 공급되는 동안 2, 3 bisphosphoglycerate가 제거된다.
적혈구에는 1, 3 bisphosphoglycerate (신진 대사에 의해 생성 된)를 2, 3 bisphosphoglycerate로 전환시켜 4-5mM의 농도에 도달하여 헤모글로빈이 l 조직 내의 산소.
조직에 도착한 헤모글로빈은 방출 된 상태 (산소에 결합되어 있음)이지만 조직 근처에서는 카르 복 실화되어 신장 된 상태로 넘어갑니다.이 상태에있는 단백질은 산소와 결합하는 경향이 낮기 때문에, 방출 된 상태와 비교하여, 헤모글로빈은 조직으로 산소를 방출한다; 또한, 물과 이산화탄소 사이의 반응으로 인해 H + 이온이 생성되고 보어 효과로 인해 산소가 더 많이 생성됩니다.
이산화탄소 는 원형질막을 통과하는 적혈구로 확산된다. 적혈구가 혈액의 약 40 %를 구성한다고 가정 할 때 조직에서 확산되는 이산화탄소의 40 %만이 이산화탄소의 약 40 %를 차지할 것으로 예상해야합니다. 실제로 이산화탄소의 90 %는 적혈구에 들어갑니다. 탄산의 이산화탄소가 적 으면 적혈구의 이산화탄소가 일정한 농도로 낮아 진입 속도가 빠르다.

적혈구가 조직에 도달 할 때 발생하는 또 다른 현상은 다음과 같습니다 : 기울기에 의해 HCO3- (이산화탄소 유도체)가 적혈구에서 나오고 음전하의 출구 균형을 맞추기 위해 삼투압의 증가를 결정하는 염화물의 유입 :이 변화의 균형을 맞추기 위해 적혈구 팽창을 일으키는 물의 침투가 있습니다 (햄버거 효과). 반대 현상은 적혈구가 폐의 폐포에 도달 할 때 발생합니다 : 적혈구의 수축이 발생합니다 (HALDANE 효과). 따라서 정맥혈 (폐에 직접 작용하는 정맥혈)은 동맥혈보다 둥글다.


